
- Строение периферического отдела вкусовой системы
- Функциональные типы вкусовых клеток
- Принцип работы рецепторов
- Афферентная нейропередача во вкусовых клетках
- Ярослав Андреев, Юлия Логашина, Ксения Лубова, Александр Василевский, Сергей Козлов «Природа» №12, 2016
- Как победить боль?
- Рецепторы температуры и химических раздражителей
- Роман Романов «Природа» №8, 2014
- Об авторе
- Рецепторы кислоты
Строение периферического отдела вкусовой системы


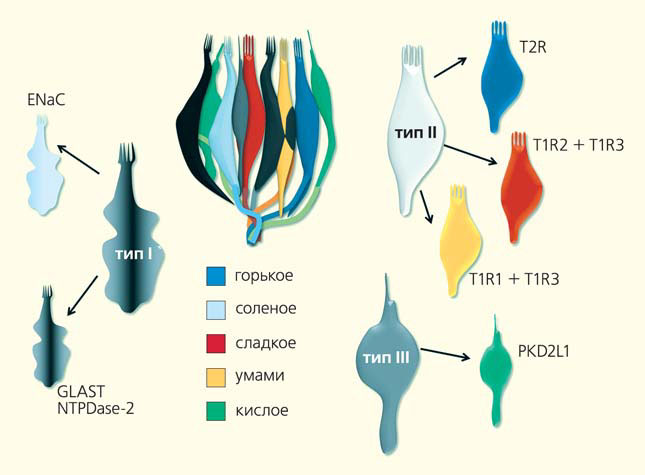
Этот отдел включает более 5000 особых образований — сосочков разной формы: грибовидных (они находятся в передней части языка), желобовидных (в его корневой доли) и листовидных (по бокам). Уже на этом уровне проявляется определенная, хотя и неполная, специализация. Так, основная информация о сладком приходит в мозг прежде всего от грибовидных сосочков, тогда как о горьком — от желобовидных, а о кислом и соленом — от листовидных. Вкусовые сосочки состоят из почек (в общей сложности их во рту около 10 тыс.) — ассоциатов из 50–100 выстроенных подобно долькам апельсина специализированных клеток вытянутой формы (рис. 2). Они обладают многими свойствами нейронов, такими как способность к генерации потенциалов действия (волн возбуждения, возникающих, как правило, при деполяризации мембраны, изменении ее потенциала в сторону положительных значений) и образование синапсов (мест контакта между двумя клетками). Но, несмотря на наличие признаков электровозбудимых тканей, вкусовые, как и сенсорные клетки органов слуха и равновесия, развиваются из эпителия. Сама вкусовая почка не достигает поверхности слизистой оболочки языка, в полость рта выходит только пора, через которую растворенные в слюне вещества контактируют со вкусовыми клетками. Их наружная часть представлена микроворсинками с расположенными на них рецепторами — главными участниками процессов распознавания, усиления и преобразования пищевых сигналов. Рецепторы микроворсинок при связывании своих лигандов (веществ, специфически соединяющихся с рецептором) запускают каскад биохимических реакций во вкусовой клетке, которая в ответ высвобождает нейромедиатор (химическое вещество, передающее электрический сигнал на нервное волокно), стимулирующий вкусовой нерв. И в итоге в мозг уходят электрические импульсы, несущие информацию об интенсивности и разновидности воздействия.
Среди вкусовых клеток выделяют как минимум три типа: темные (I), так как они интенсивно окрашиваются на ультратонких срезах, светлые (II) и промежуточные (III). Апикальная часть, непосредственно контактирующая с ротовой полостью, обычно богата микроворсинками (исключение составляют клетки III типа, у них только одна микроворсинка) и несет метаботропные рецепторы (вызывающие изменения в клеточном метаболизме после связывания со своими лигандами), сопряженные с G-белками, ионные каналы и другие сигнальные белки. Базальная часть, не имеющая доступа ко вкусовым веществам, отвечает за связь с афферентными и эфферентными (проводящими возбуждение от органов и тканей в центральную нервную систему и наоборот) нервными окончаниями и, возможно, с другими рецепторными клетками вкусовой почки. Апикальные части всех трех типов клеток соединены друг с другом окружающими их плотными контактами — белковыми образованиями, которые выглядят как пояс из переплетающихся нитей и служат барьером для диффузии вкусовых веществ. Средняя продолжительность жизни вкусовой клетки не превышает 10 дней.
Функциональные типы вкусовых клеток
Принцип работы рецепторов
Стимулами для разных рецепторов могут служить свет, механическая деформация, химические вещества, изменения температуры, а также изменения электрического и магнитного поля. В рецепторных клетках (будь то непростредственно нервные окончания или специализированные клетки) соответствующий сигнал изменяет конформацию чувствительных молекул-клеточных рецепторов, что приводит к изменению активности мембранных ионных рецепторов и изменению мембранного потенциала клетки. Если воспринимающей клеткой является непосредственно нервное окончание (так называемые первичные рецепторы), то обычно происходит деполяризация мембраны с последующей генерацией нервного импульса. Специализированные рецепторные клетки вторичных рецепторов могут как де-, так и гиперполяризоваться. В последнем случае изменение мембранного потенциала ведет к уменьшению секреции тормозного медиатора, действующего на нервное окончание и, в конечном счете, все равно к генерации нервного импульса. Такой механизм реализован, в частности, в чувствительных элементах сетчатки.
В качестве клеточных рецепторных молекул могут выступать либо механо-, термо- и хемочувствительные ионные каналы, либо специализированные G-белки (как в клетках сетчатки). В первом случае открытие каналов непосредственно изменяет мембранный потенциал (механочувствительные каналы в тельцах Пачини), во втором случае запускается каскад внутриклеточных реакций трансдукции сигнала, что ведет в конечном счете к открытию каналов и изменению потенциала на мембране.
Афферентная нейропередача во вкусовых клетках
Как же вкусовые клетки разных типов передают информацию на нервные окончания? Обычно нейропередача происходит благодаря синаптическим пузырькам, наполненным афферентным нейромедиатором. При слиянии с клеточной мембраной (кальций-зависимом экзоцитозе) они высвобождают свое содержимое в межклеточное пространство, возбуждая близлежащее нервное окончание. За открытие молекулярных основ везикулярного транспорта Джеймсу Ротману, Ренди Шекману и Томасу Зюдофу в 2013 г. была присуждена Нобелевская премия по физиологии или медицине. Однако только в клетках III типа, распознающих кислое, имеются синаптические пузырьки и другие атрибуты классических химических синапсов, включая белки комплекса SNARE (они осуществляют экзоцитоз) и потенциал-зависимые кальциевые каналы (благодаря которым процесс экзоцитоза запускается). Ранее, кстати, из-за того что образование синапсов с афферентным нервным окончанием присуще лишь клеткам III типа, они ошибочно рассматривались как единственные истинно рецепторные. Кислые вещества вызывают открытие ионных каналов PKD2L1 и других протон-чувствительных каналов клеток III типа. Входящий ток (обусловленный поступлением в клетку ионов) приводит к деполяризации мембраны выше критического уровня и к генерации серии потенциалов действия, частота которых зависит от силы тока, пропорционального, в свою очередь, концентрации протонов во вкусовой поре. Это влечет за собой активацию потенциал-зависимых кальциевых каналов и вход кальция в клетку. Повышение его концентрации в цитозоле запускает экзоцитоз синаптических пузырьков и высвобождение нейромедиатора серотонина на вкусовое нервное окончание. Таким образом, нейропередача клетками III типа осуществляется вполне традиционным способом (рис. 4).


При вкусовой стимуляции в клетках II типа можно предположить следующую цепь событий (рис. 6). Вкусовые молекулы связываются с G-белковыми рецепторами T1R и T2R, расположенными в апикальной части клетки. Это приводит к активации G-белков (в частности, гастдуцина), а затем — фосфолипазы С β2 и вызывает мобилизацию (увеличение концентрации) внутриклеточного кальция (скорее всего, при участии IP3-рецептора эндоплазматической сети). Он открывает Ca2+-зависимый ионный канал TRPM5, чем способствует осцилляционной деполяризации мембраны и генерации серии потенциалов действия. Это стимулирует потенциал-зависимые ионные каналы CALHM1, проницаемые для АТФ, и ведет к его высвобождению из клетки. При этом генерация серии потенциалов действия позволяет закодировать информацию об интенсивности вкусового воздействия. Секретированный АТФ активирует P2X2/P2X3 рецепторы афферентных нервных окончаний, и сигнал о пищевом воздействии уходит в мозг. Роль вкусовой клетки I типа в основном, по-видимому, состоит в гидролизе АТФ, высвобождаемого на нервное окончание, с помощью уже упомянутой эктоапиразы 2.

Многие, несомненно, задаются вопросом: зачем изучать механизмы вкусового восприятия? Действительно ли нам необходимо знать, какие клетки и как в нем участвуют? Оказывается, значительное число людей страдает расстройством вкуса. У большинства оно развивается после травмы головы или инфекции верхних дыхательных путей и среднего уха; после лучевой терапии (при лечении онкологических заболеваний головы и шеи); в результате действия определенных химикатов (инсектицидов и некоторых лекарств, включая антибиотики и антигистаминные средства) и как последствие некоторых видов хирургии уха, носа и горла. Опасны ли расстройства вкуса? Да, так как они ослабляют или вовсе устраняют врожденную систему предупреждения об испорченной пище, а иногда и об аллергенах. При ухудшении чувства вкуса мы можем начать есть слишком мало / много и терять / набирать вес. Потеря вкуса может вынудить нас употреблять чрезмерные количества сахара или соли, чтобы улучшить вкус пищи, а для людей с сахарным диабетом или высоким кровяным давлением это очень рискованно. В тяжелых случаях потеря вкуса может даже привести к депрессии. Поэтому знания о механизмах его восприятия необходимы для изучения причин, приводящих к отклонениям, и поиска способов их коррекции. А это — важная научная и медицинская задача.
Литература
1. Благутина В. В. Анатомия вкуса // Химия и жизнь. 2010. № 10. C. 34–38.
2. Lindemann B. Receptors and transduction in taste // Nature. 2001. V. 413. P. 219–225.
3. Margolskee R. F. Molecular mechanisms of bitter and sweet taste transduction // J. Biol. Chem. 2002. V. 277. P. 1–4.
4. Zhang Y., Hoon M. A., Chandrashekar J. et al. Coding of sweet, bitter, and umami tastes, different receptor cells sharing similar signaling pathways // Cell. 2003. V. 112. P. 293–301.
5. Huang A. L., Chen X., Hoon M. A. et al. The cells and logic for mammalian sour taste detection // Nature. 2006. V. 442. P. 934–938.
6. Bartel D. L., Sullivan S. L., Lavoie E. G. et al. Nucleoside triphosphate diphosphohydrolase-2 is the Ecto-ATPase of type I cells in taste buds // J. Comp. Neurol. 2006. V. 497. P. 1–12.
7. Lawton D. M., Furness D. N., Lindemann B. et al. Localization of the glutamate-aspartate transporter, GLAST, in rat taste buds // Eur. J. Neurosci. 2000. V. 12. P. 3163–3671.
8. Ralevic V., Burnstock G. Receptors for purines and pyrimidines // Pharmacol. Rev. 1998. V. 50. P. 413–492.
9. Finger T. E., Danilova V., Barrows J. et al. ATP signaling is crucial for communication from taste buds to gustatory nerves // Science. 2005. V. 310. P. 1495–1499.
10. Romanov R. A., Kolesnikov S. S. Electrophysiologicaly identified subpopulations of taste bud cells // Neurosci. Lett. 2006. V. 395. P. 249–254.
11. Romanov R. A., Rogachevskaja O. A., Bystrova M. F. et al. Afferent neurotransmission mediated by hemichannels in mammalian taste cells // EMBO J. 2007. V. 26. P. 657–667.
12. Bystrova M. F., Romanov R. A., Rogachevskaya O. A. et al. Functional expression of the extracellular calcium-sensing receptor in mouse taste cells // J. Cell Sci. 2010. V. 123. P. 972–982.
13. Lazarowski E. R., Boucher R. C., Harden T. K. Mechanisms of release of nucleotides and integration of their action as P2X- and P2Y-receptor activating molecules // Mol. Pharmacol. 2003. V. 64. P. 785–795.
14. Romanov R. A., Bystrova M. F., Rogachevskaya O. A. et al. The ATP permeability of pannexin 1 channels in a heterologous system and in mammalian taste cells is dispensable // J. Cell Sci. 2012. V. 125. P. 5514–5523.
15. Romanov R. A., Rogachevskaja O. A., Khokhlov A. A. et al. Voltage-dependence of ATP secretion in mammalian taste cells // J. Gen. Physiol. 2008. V. 132. P. 731–744.
16. Siebert A. P., Ma Z., Grevet J. D. et al. Structural and functional similarities of calcium homeostasis modulator 1 (CALHM1) ion channel with connexins, pannexins, and innexins // J. Biol. Chem. 2013. V. 288. P. 6140–6153.
17. Taruno A., Vingtdeux V., Ohmoto M. et al. CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes // Nature. 2013. V. 495. P. 223–226.
18. Murata Y., Yasuo T., Yoshida R. et al. Action potential-enhanced ATP release from taste cells through hemichannels // J. Neurophysiol. 2010. V. 104. P. 896–901.
Ярослав Андреев, Юлия Логашина, Ксения Лубова, Александр Василевский, Сергей Козлов
«Природа» №12, 2016

Говорят, что жизнь — это боль. Хотя в этой фразе содержится нечто негативное, связанное с неприятными ощущениями, переживаниями или даже тяжелыми страданиями, не стоит забывать, что боль (ноцицепция) предупреждает нас об опасности — сигнализирует о нарушениях в организме, который немедленно принимается их устранять. Вместе с тем существует и боль, которая приносит только мучения.
Основная причина появления такой боли — сбои в передаче болевых сигналов (нервных импульсов) от чувствительных нейронов к головному мозгу, который и формирует неприятные ощущения. Когда воздействие неопасных стимулов распознающие нейроны расценивают как опасное, развивается состояние, которое называется гиперчувствительностью. И это не всегда плохо, так как в нужный момент она играет важную роль в процессе выздоровления и восстановления организма. Однако бывает и так, что реального повода нет, а гиперчувствительность ведет к изнурительной хронической боли. В таком случае самые обычные безобидные стимулы (легкое прикосновение или тепло) вызывают аллодинию (от греч. άλλος — другой и οδύνη — мучение), а болезненные стимулы — боль еще большей интенсивности, гиперальгезию (от греч. ὑπέρ — сверх- и ἄλγος — боль). Часто аномально интенсивная и нередко хроническая боль, которая изматывает и физиологически, и психологически, а также затрудняет выздоровление, возникает в результате таких заболеваний, как артрит, опоясывающий лишай, СПИД, рак костей и др.
Прежде чем винить в аномалиях чувствительные нейроны (ноцицепторы), которые воспринимают, анализируют и передают болевые сигналы, разберемся, как они работают в здоровом организме и что происходит при патологиях.
Как победить боль?
Так что же делать, если мы испытываем боль? Если это боль острая или хроническая, терпеть ее нельзя, и необходимо использовать обезболивающие средства, чтобы вернуть нашу систему ноцицепции в нормальное состояние, а себя — к жизни в самом прямом смысле этого слова. В настоящее время для обезболивания применяется множество лекарственных препаратов различных фармакологических групп. Основное место в этом ряду занимают нестероидные противовоспалительные средства (НПВС), антиконвульсанты и антидепрессанты, а также наркотические анальгетики (морфин и другие опиаты и опиоиды). Имеющиеся в настоящее время анальгетические средства влияют главным образом на пути передачи и распространения боли. Для специфичного регулирования рецепторов боли, описанных выше, пока препаратов на рынке лекарств нет.
Рецепторы чувствительных нейронов представляют собой заманчивую, но сложную мишень для создания лекарств. Препараты, если они обладают хорошей селективностью к этим рецепторам, будут приняты потребителями с большой радостью, так как почти все современные средства ограничены в применении из-за побочных эффектов. Работы по поиску селективных препаратов ведутся, в том числе и в нашей стране, и при благоприятном стечении обстоятельств такие лекарства уже скоро смогут появиться в аптеках. Долгих вам лет жизни без боли!
Работа выполнена при поддержке Российского научного фонда (проект № 14-24-00118).
Рецепторы температуры и химических раздражителей

TRP-рецепторы — тетрамеры (рис. 3), то есть образованы четырьмя полипептидными цепочками. При этом могут собираться как гомомеры, то есть рецепторы, сформированные одинаковыми цепочками (например, TRPV1 или TRPV3, описанные выше), так и гетеромеры из разных цепей. Гетеромерные рецепторы (например, построенные из цепочек TRPV1 и TRPV3) обладают различной чувствительностью к тепловым стимулам, пороговая температура их активации лежит между значениями, пороговыми для гомомерных рецепторов.
Воздействие на рецепторы чувствительных нейронов, участвующие в восприятии температуры, с помощью лекарственных средств приводит к облегчению боли и воспаления. Именно так, не зная о молекулярных мишенях, народная медицина в разное время применяла настойки перца (TRPV1), горчицы (TRPA1), мяты (TRPM8) и гвоздики (TRPV3) для лечения ряда воспалительных заболеваний.
Роман Романов
«Природа» №8, 2014

Об авторе
Роман Александрович Романов — доктор биологических наук, старший научный сотрудник лаборатории молекулярной физиологии клетки Института биофизики клетки РАН (Пущино, Московская обл.). Научные интересы связаны с механизмами афферентной нейропередачи во вкусовых клетках.
С чего начинается то удовольствие, которое мы неизменно испытываем, отправляя в рот ложечку любимого блюда? Первый этап вкусового восприятия — возбуждение специализированных сенсорных клеток языка в ответ на появление еды. Их основополагающая функция заключается в распознавании растворенных в жидкой среде полости рта химических молекул и в передаче информации об их концентрации и модальности (разновидности) для дальнейшего анализа в соответствующих структурах мозга. Вкусовая система состоит из трех отделов — периферического, представленного сенсорными клетками, проводникового (нервные волокна, идущие к головному мозгу) и центрального, или коркового (нейроны вкусовой зоны коры больших полушарий) (рис. 1). Распознавание вкусовых веществ происходит в периферическом отделе, а передача полученной информации в центр обработки — корковую зону — осуществляется проводниковым отделом. Пищевые стимулы очень многообразны, от простых ионов (Na+, H+) массой в несколько дальтон до сложных молекул, таких как углеводы, алкалоиды, белки, масса которых может заметно превышать 1 кДа. Большинство вкусовых оттенков, которые ассоциированы с ароматом (например, кофе, корицы, вина), связаны с активацией еще и обонятельных рецепторных клеток носовой полости. Вообще чувство вкуса связано с раздражением не только химических и обонятельных, но и механических (вяжущее ощущение), температурных (мята, ментол) и даже болевых (перец чили) рецепторов. Непосредственно же с помощью вкусовой системы человек распознает пять категорий: сладкое (природные углеводы), кислое (лимонная и уксусная кислоты), соленое (поваренная соль), горькое (никотин, хинин) и умами (L-глутамат и некоторые другие аминокислоты, мелкие пептиды и пурины). В ходе эволюции у млекопитающих (речь о них, а значит, и о нас с вами, пойдет в этой статье) развилась высокая чувствительность к горьким веществам (так как они нередко ядовиты) и к сладким — часто высокоэнергетическим продуктам.

Рецепторы кислоты
В отличие от TRP-рецепторов, P2X-рецепторы и ASIC являются тримерами (рис. 3), т.е. собраны из трех полипептидных цепочек. Но точно так же эти рецепторы могут быть гомомерами и гетеромерами, что увеличивает их разнообразие и спектр выполняемых функций.