
- Серотониновый рецептор 5-HT1A
- Распределение в организме
- Сигнальный путь MAPK/ERK
- Влияние активации 5-HT₁A-рецепторов на активность MEK/ERK каскада
- Эффект на клетки гиппокампа
- Реакция нейрональных клеток из разных областей
- Факторы, влияющие на ответ MEK/ERK-сигнального каскада
- Выводы
- Регуляция активности ERK с использованием 5-HT₁A-рецепторов в мозге
- Гипоталамус
- Гиппокамп
- Кора больших полушарий
- Фронтальная кора
- Префронтальная кора
- Ядра шва
- Влияние селективной активации
- Сигнальный путь PI3K/Akt
- Эффекты 5-HT1A-рецептора в центральной нервной системе
- Эндокринные эффекты 5-HT1A-рецептора
- Взаимодействия рецептора с другими белками
- Палочки и колбочки — что это?
- Симптоматика заболеваний палочек и колбочек
- Диагностика и лечение заболеваний палочек и колбочек
- Код опасности
- Как мы воспринимаем горечь
- Супердегустаторы, дегустаторы и недегустаторы
- Полиморфизмы и здоровье
- TAS2R и пищевое поведение
- Заключение
Серотониновый рецептор 5-HT1A
5-гидрокситриптаминовый (серотониновый) рецептор 1А, связанный с G-белком, 5-HT-1A, 5-HT1A, 5HT1a, ADRB2RL1, ADRBRL1, G-21, PFMCD, 5-гидрокситриптаминовый рецептор 1A
- Внешние идентификаторы:
- OMIM: 109760
- MGI: 96273
- HomoloGene: 20148
- GeneCards: HTR1A
Расположение гена:
| Вид | Хр.Локус | Начало | Конец |
|---|---|---|---|
| Человек | |||
| Мышь |
Паттерн экспрессии РНК:
- Bgee:
- Человек: middle temporal gyrus, Brodmann area 46, superior vestibular nucleus, hippocampus proper
- Мышь: ventral nuclear group, ventral posterolateral nucleus, ventral posteromedial nucleus, thalamic reticular nucleus, Region I of hippocampus proper, hippocampus proper
Дополнительные справочные данные:
- BioGPS
Ортологи:
| Вид | Entrez | Ensembl | UniProt | RefSeq (мРНК) | RefSeq (белок) | Локус (UCSC) |
|---|---|---|---|---|---|---|
| Человек | Chr 5: 63.96 – 63.96 Mb | |||||
| Мышь | Chr 13: 105.58 – 105.58 Mb |
Информация в Викиданных
Серотониновый рецептор подтипа 5-HT1A является белком (гликопротеином), состоящим из 422 аминокислот (молекулярная масса 46107 дальтон). У него есть семь трансмембранных и α-спиральных доменов, а также активный сайт для связывания с лигандами, включая серотонин. Гликозируется в позициях 10, 11 и 24 по аспарагину, а также связывается с убиквитином в позиции лизина 334.
Распределение в организме
Серотониновый рецептор 5-HT1A связан с Gi и участвует в модуляции активности калиевых каналов и электрической активности клеток. Его активация также активирует фосфолипазу С, возбуждающую образование вторичных мессенджеров like диацилглицерина (DAG) и инозитолтрифосфата (IP3), а также протеинкиназы C, провоцирующие фосфорилирование белков нисходящего эффекторного каскада.
Взаимодействие с системой синтазы оксида азота и оксидом азота также оказывает влияние на активность 5-HT1A-рецепторов.
Изучение сигнальных каскадов 5-HT1A-рецептора предполагает некоторые сложности.
Несмотря на то, что связь 5-HT₁A-рецепторов с опосредованными Giα / Goα изменениями активности аденилатциклазы, концентрации внутриклеточного цАМФ и активности протеинкиназы А и с опосредованными Giβγ / Goβγ изменениями активности GIRK и тока ионов калия уже хорошо установлена, функция этих двух канонических сигнальных механизмов и их роль в опосредованной 5-HT₁A-рецепторами регуляции активности нейронов всё ещё не изучена достаточно полно.
Сложность механизмов внутриклеточной передачи сигнала в нейронах требует комбинированных фармакологических, биохимических и молекулярно-биологических подходов к их изучению для детальной характеристики каждого компонента внутриклеточного сигнального каскада и его роли и места в общей схеме. А это сильно ограничивает возможности изучения этих каскадов in vivo и возможность охарактеризовать их влияние на функции нейронов в их естественной среде, на функцию мозга в целом и на поведение живых организмов как целого.
Помимо этого, в работе 5-HT₁A-рецепторов могут играть роль также дополнительные, неканонические сигнальные каскады. Среди них в последнее время наибольшее внимание привлекают сигнальные каскады, работа которых традиционно ассоциировалась с активацией рецепторов для факторов роста, и которые также оказались связаны с работой 5-HT₁A-рецепторов.
Сигнальный путь MAPK/ERK
Способность 5-HT₁A-рецепторов активировать ERK1/2 при помощи их фосфорилирования была впервые обнаружена в клетках, отличных от нейронов, но также экспрессирующих 5-HT₁A-рецепторы. Это воздействие 5-HT₁A-рецепторов на активность ERK1/2 чувствительно к ингибированию Gi-белков коклюшным токсином.
Это заставляет предполагать, что в активации ERK-сигнального пути при стимуляции 5-HT₁A-рецепторов принимает участие G-белок-связанный механизм. Так же, как и при вызванной факторами роста активации ERK, вызванная активацией 5-HT₁A-рецепторов активация ERK опосредуется активацией малых ГТФ-аз Ras и Raf и активацией через них MEK.
Этот сигнальный каскад требует кальмодулин-зависимого эндоцитоза рецепторов как промежуточного этапа. Кроме того, активация ERK1/2 при активации 5-HT₁A-рецепторов в ненейронных клетках может также опосредоваться фосфатидилинозитол-3-киназой (PI3K) и фосфатидилхолин-специфичной фосфолипазой С (PLC). Этот эффекторный каскад также является G-белок-зависимым.
Однако детали процесса передачи сигнала от G-белок-зависимой фосфатидилинозитол-3-киназы (PI3K) к ERK в результате активации 5-HT₁A-рецепторов ещё до конца не понятны.
Влияние активации 5-HT₁A-рецепторов на активность MEK/ERK каскада
Несмотря на то, что в ненейронных клеточных системах активация 5-HT₁A-рецепторов систематически и закономерно приводит к активации сигнального каскада MEK/ERK, эффекты активации 5-HT₁A-рецепторов на активность MEK/ERK каскада и в частности на активность ERK1/2 в клетках нейронального происхождения вариабельны в зависимости от места происхождения и степени дифференцировки нейронов.
Эффект на клетки гиппокампа
В высокодифференцированных HN2-5 клетках, происходящих из гиппокампа, агонисты 5-HT₁A-рецепторов вызывают усиление фосфорилирования ERK1/2 и повышение её активности. Показано, что этот эффект зависит от активности малых ГТФ-аз Ras и Raf, активации MEK и мобилизации внутриклеточного кальция. Однако этот эффект активации 5-HT₁A-рецепторов не обнаруживается в первичной культуре низкодифференцированных гиппокампальных нейронов или в ромбоэнцефалических нейронах эмбриона.
Реакция нейрональных клеток из разных областей
Для низкодифференцированных нейронов активация 5-HT₁A-рецепторов нейтральна в отношении активности ERK1/2, не влияет на неё. А в высокодифференцированных клетках из ядер шва активация 5-HT₁A-рецепторов приводит к противоположному эффекту — ингибированию активности MEK и снижению фосфорилирования и активности ERK.
Факторы, влияющие на ответ MEK/ERK-сигнального каскада
Многие факторы влияют на реакцию MEK/ERK-сигнального каскада на активацию 5-HT₁A-рецепторов. Для проявления активирующего действия 5-HT₁A-рецепторов на активность ERK необходима высокая плотность 5-HT1A-рецепторов на поверхности клеток, что чаще встречается у зрелых, высокодифференцированных клеток.
Тип 5-HT₁A-рецепторов (ауторецепторы или гетерорецепторы), их связь с Gi / Go ингибиторными белками, а также доступность этих белков в клетках, оказывают влияние на ERK-ответ на активацию 5-HT₁A-рецепторов.
Выводы
Опосредуемая 5-HT₁A-рецепторами регуляция активности MEK/ERK-сигнального пути высоко селективна и специфична в зависимости от области происхождения нейронов, их возраста и степени дифференцировки. Это важно учитывать, так как в мозгу существует множество разнообразных типов нейронов, экспрессирующих 5-HT₁A-рецепторы, и различные области с разными функциями, содержащие такие нейроны.
Регуляция активности ERK с использованием 5-HT₁A-рецепторов в мозге
В нескольких исследованиях было убедительно показано, что активация MEK/ERK-сигнального пути при активации 5-HT₁A-рецепторов не является универсальной реакцией для любых нейронов мозга.
Гипоталамус
В частности, в ряде исследований было показано, что 5-HT₁A-агонисты быстро, но кратковременно повышают фосфорилирование ERK в гипоталамусе, и что этот эффект, по всей вероятности, является промежуточным этапом в вызываемой 5-HT₁A-агонистами индукции биосинтеза и высвобождения окситоцина, АКТГ и пролактина.
Гиппокамп
В контрасте с этим, 5-HT₁A-агонисты понижают фосфорилирование ERK и активность ERK в гиппокампе. И хотя значение факта этой отрицательной привязки активности ERK к активности 5-HT₁A-рецепторов в гиппокампе точно не известно, но сама по себе ERK известна как критически важный регулятор процессов мышления и памяти и как важный медиатор синаптической пластичности.
Кора больших полушарий
Вероятно, ингибирование 5-HT₁A-рецепторами активности ERK в гиппокампе может играть роль в опосредуемых 5-HT₁A-рецепторами изменениях синаптической пластичности или в объяснении вызываемых активацией 5-HT₁A-рецепторов нарушений когнитивных функций. Находки, характеризующие регуляцию 5-HT₁A-рецепторами фосфорилирования ERK в других областях мозга, таких, как кора больших полушарий, миндалевидное тело, ядра шва, значительно более противоречивы.
Фронтальная кора
Так, например, по некоторым данным, во фронтальной коре 5-HT₁A-агонисты повышают фосфорилирование ERK, а по другим данным, там они не влияют на степень фосфорилирования ERK.
Префронтальная кора
В префронтальной коре активация 5-HT₁A-рецепторов или NMDA-рецепторов по отдельности никак не влияет на активность ERK, однако одновременная активация обоих типов рецепторов приводит к снижению фосфорилирования ERK и к понижению её активности.
Ядра шва
По контрасту с исследованиями, которые показали, что в изолированных высокодифференцированных нейронах ядер шва активация 5-HT₁A-рецепторов приводит к снижению активности ERK, показано, что введение 5-HT₁A-агонистов в живой организм приводит к кратковременному увеличению активности ERK в дорсальной части ядер шва — эффект, полностью противоположный наблюдаемому в культуре тех же самых клеток.
Влияние селективной активации
Кроме того, селективная активация 5-HT₁A-ауторецепторов ядер шва (а их там очень много) может также косвенно, опосредованно влиять на активность MEK/ERK-сигнального пути в мозгу через регуляцию высвобождения серотонина в других областях мозга.
Для более чёткого понимания взаимосвязи между активностью 5-HT₁A-рецепторов и активностью сигнального каскада MEK/ERK в коре больших полушарий нужны более детальные исследования этих связей в различных областях коры и различных типах нейронов коры.
Таким образом, регуляция 5-HT₁A-рецепторами активности ERK в мозгу очень сложна и разнообразна и отличается в разных областях мозга и для разных типов нейронов. Для выяснения деталей специфической регуляции 5-HT1A-рецепторами активности ERK в разных областях мозга необходимы дальнейшие эксперименты с локальной активацией 5-HT₁A-рецепторов в тех или иных областях мозга и опыты с трансгенными мышами с пространственными и временными модификациями 5-HT₁A-рецепторов.
Учитывая выраженное влияние 5-HT₁A-рецепторов на регуляцию настроения, уровня тревожности, память и когнитивные функции, и наличие данных, показывающих выраженное регионарно-специфичное и типоспецифичное для разных типов нейронов влияние 5-HT₁A-рецепторов на активность ERK, роль ERK в опосредовании поведенческих эффектов 5-HT₁A-рецепторов и в регуляции активности нейронов заслуживает дальнейшего изучения, так же как и возможный терапевтический потенциал непосредственной фармакологической модуляции активности этого важного регулируемого 5-HT₁A-рецепторами сигнального каскада на более низких уровнях (на уровне тех или иных внутриклеточных звеньев, а не на уровне 5-HT₁A-рецептора).
Сигнальный путь PI3K/Akt
Серотониновые рецепторы подтипа 5-HT₁A также способны активировать другой сигнальный путь, обычно активируемый различными факторами роста, так называемый сигнальный путь PI3K / Akt. Когда тирозинкиназные рецепторы активируются факторами роста, они рекрутируют (активируют, фосфорилируя по тирозину) PI3K. Активированная PI3K, в свою очередь, активирует фосфоинозитид-зависимую киназу (PDK). А активированная PDK фосфорилирует и активирует Akt. Белок Akt хорошо известен как регулятор клеточного роста и выживаемости и как фактор, противодействующий апоптозу и способствующий митотической активности. Активация белка Akt ростовыми факторами опосредует вызываемый инсулином рост тканей и повышает устойчивость клеток к про-апоптотическим воздействиям. В центральной нервной системе белок Akt является важнейшим эффекторным белком в сигнальных каскадах, реализующих эффект нейротрофинов и различных нейромедиаторов. Также как и ERK, белок Akt является мультиспецифичной протеинкиназой широкого назначения, которая фосфорилирует множество мишеней-субстратов, в том числе протеинкиназы нисходящего эффекторного сигнального пути PI3K / Akt, различные факторы транскрипции белков и другие регуляторные белки.
Киназа гликоген-синтетазы 3 (GSK3) — это одна из протеинкиназ, которая в основном фосфорилируется и инактивируется белком Akt, но также может фосфорилироваться и инактивироваться несколькими другими протеинкиназами, такими, как протеинкиназа C или протеинкиназа А (цАМФ-зависимая). Киназа гликоген-синтетазы 3 (GSK3) является потенциальной молекулярной мишенью при некоторых психических заболеваниях, прежде всего заболеваниях аффективной сферы. В частности, известно, что нормотимик литий является селективным ингибитором GSK3. Ингибирование GSK3 фармакологическим методом или выведение лишённой GSK3 трансгенной мыши имитирует эффект антидепрессантов и антиманиакальных средств (нормотимиков и антипсихотиков). И наоборот, нарушенная регуляция GSK3 приводит к поведенческим аномалиям у животных, напоминающим состояния мании и депрессии.
Другая интересная группа субстратов фосфорилирования белком Akt — это так называемые транскрипционные факторы Forkhead-box-O (также называемые FoxOs). Активированный в результате действия факторов роста белок Akt фосфорилирует FoxOs и инактивирует их, вызывая их экспорт из ядра клетки в цитоплазму, где они не могут влиять на транскрипцию ДНК. И у беспозвоночных (в частности, насекомых, как фруктовая мушка дрозофила), и у высших животных белки FoxOs также могут подвергаться фосфорилированию и инактивации при воздействии серотонина через 5-HT₁A-рецептор и нисходящий эффекторный PI3K / Akt-зависимый механизм. В частности, показано, что подтип FoxO3a этого белка, распространённый в мозгу, может инактивироваться при воздействии имипрамина и подвергаться даунрегуляции (понижению активности и степени экспрессии) при воздействии ионов лития. Кроме того, трансгенные мыши с дефицитом FoxO проявляют поведение, характерное для мышей, получающих антидепрессанты и/или анксиолитики.
Таким образом, регуляция активности различных белков при помощи сигнального пути PI3K / Akt играет важную роль не только в росте и развитии нейронов и в их устойчивости к апоптозу, но и в поддержании активности нейронов и в регулировании поведенческих реакций, и этот сигнальный путь, по-видимому, тоже является важным для реализации антидепрессивного и анксиолитического эффектов 5-HT₁A-рецепторов.
В клетках, отличных от нейронов, активация гетерологичных 5-HT₁A-рецепторов закономерно повышает фосфорилирование белка Akt и его активность. Подобно регуляции белка ERK, регуляция Akt при помощи 5-HT₁A-рецепторов опосредуется Gi / Go и нисходящими эффекторными каскадами PI3K и Ras. Кроме того, активация Akt при активации 5-HT₁A-рецепторов предотвращается при повышении концентрации циклического АМФ в клетке (например вызванном одновременной активацией каких-то других, стимулирующих, а не ингибирующих аденилатциклазную активность, рецепторов) и соответствующей активации протеинкиназы A. Это влияние накопления цАМФ на активность Akt предотвращается или снимается ингибированием или инактивированием протеинкиназы А. Это заставляет предполагать, что вызываемое активацией 5-HT₁A-рецепторов ингибирование активности аденилатциклазы с соответствующим снижением содержания цАМФ в клетке и активности протеинкиназы A также является одним из механизмов, при помощи которых активация 5-HT₁A-рецепторов повышает активность Akt. Это согласуется с другими данными, которые показывают, что накопление цАМФ в клетке может вызывать дефосфорилирование и инактивацию Akt за счёт активации протеинкиназы A (PKA) и PKA-зависимой активации протеин-фосфатаз, в том числе протеинфосфатазы 1.
Было показано, что агонисты 5-HT₁A-рецепторов закономерно повышают фосфорилирование и активность Akt в нейронах разного происхождения, включая происходящие из гиппокампа HN2-5 нейрональные клетки разной степени дифференцировки и зрелости, первичные низкодифференцированные гиппокампальные нейроны, первичные ромбоэнцефалические нейроны плода. Так же, как и в клетках не-нейронного происхождения, вызываемая активацией 5-HT₁A-рецепторов активация белка Akt является G-белок-зависимым процессом, чувствительным к ингибированию Gi и повышению активности аденилатциклазы коклюшным токсином и к концентрации цАМФ и опосредуемой ею активности протеинкиназы A. Этот процесс активации белка Akt под влиянием активации 5-HT₁A-рецепторов медиируется повышением активности PI3K, а также угнетением активности аденилатциклазы, снижением концентрации циклического АМФ и угнетением активности протеинкиназы А. Таким образом, опосредуемая отчасти PI3K, отчасти «каноническим» аденилатциклазным механизмом регуляция белка Akt и нижележащих звеньев сигнального каскада PI3K / Akt, вероятно, также является одним из важных механизмов, которыми опосредуются поведенческие и физиологические эффекты активации 5-HT₁A-рецепторов.
Прямые доказательства регуляции белка Akt активностью 5-HT₁A-рецепторов в мозге млекопитающих в нормальных физиологических условиях пока не получены. Однако существует немало косвенных доказательств участия 5-HT₁A-рецепторов в регуляции активности белка Akt и в целом сигнального каскада PI3K / Akt. Так, например, введение мыши 5-HT₁A-агониста 8-OH-DPAT значительно повышало фосфорилирование N-терминальных остатков серина у белка GSK3 — а белок GSK3 является одной из основных мишеней-субстратов белка Akt. Этот эффект наблюдался в том числе и в мозге мыши, в нескольких областях. Более того, было показано, что вызванное серотонином повышение фосфорилирования GSK3 опосредуется именно через 5-HT₁A-рецепторы (не через другие типы серотониновых рецепторов) и блокируется антагонистами 5-HT₁A-рецепторов. Кроме того, было показано, что в мозге мыши повышение уровня серотонина (при введении антидепрессантов, лития, L-триптофана) приводит к повышению фосфорилирования и активности белка Akt — эффект, который предотвращался введением в желудочки мозга ингибитора PI3K. Однако ни одно из этих исследований не предоставляет прямых доказательств того, что 5-HT₁A-рецепторы регулируют активность белка Akt в нормальных физиологических условиях в мозге млекопитающих. Также неизвестно, является ли влияние 5-HT₁A-рецепторов на активность белка Akt регионарно-специфичным и/или типоспецифичным для типа нейронов, как это имеет место быть с белками ERK. Дополнительные исследования также необходимы и для того, чтобы окончательно определить, какую роль играет регуляция PI3K / Akt сигнального пути 5-HT₁A-рецепторами в общей картине действия 5-HT₁A-рецепторов на нейрональную активность и на поведение животных.
Взятые вместе, все эти данные свидетельствуют о том, что 5-HT₁A-рецепторы сопряжены с активацией не только канонического Gi / Go-опосредованного сигнального каскада, включающего аденилатциклазу, цАМФ и протеинкиназу A, а также G-белок-зависимые каналы ионов калия (GIRK), но и с активацией добавочных сигнальных каскадов, обычно ассоциируемых с факторами роста клеток и с модуляцией нейрональной пластичности, роста и развития клеток и с их устойчивостью к апоптозу — в частности, с активацией ERK-сигнального пути и сигнального пути PI3K / Akt. Для детального изучения того, каким образом работа этих сигнальных каскадов зависит от типа клеток и от регионарной специфичности в разных областях мозга, и каким образом они медиируют разнообразные физиологические и поведенческие аспекты действия 5-HT₁A-рецепторов, требуются дополнительные исследования. Лучшее понимание механизмов передачи сигнала от 5-HT₁A-рецептора внутрь клетки может способствовать обнаружению новых мишеней для лекарств, и в конечном итоге — разработке новых лекарств (в частности, анксиолитиков и антидепрессантов) для лечения патологических состояний, ассоциирующихся с аномальным функционированием 5-HT₁A-рецепторов или их внутриклеточных каскадов (а это такие состояния, как тревожность, депрессия).
Эффекты 5-HT1A-рецептора в центральной нервной системе
Другие наблюдавшиеся в научных исследованиях результаты активации 5-HT₁A-рецепторов включают в себя:
Эндокринные эффекты 5-HT1A-рецептора
Достаточно высокие дозы агонистов 5-HT1A-рецепторов также могут эффективно обходить вызываемое пресинаптическими 5-HT₁A-ауторецепторами ингибирование высвобождения серотонина и увеличивать серотонинергическую нейропередачу, непосредственно стимулируя постсинаптические (эффекторные) 5-HT₁A-рецепторы, вместо серотонина. Однако, по контрасту с СВА, агонисты 5-HT₁A-рецепторов не в состоянии обойти ингибирующие эффекты пресинаптических 5-HT₁A-рецепторов, расположенных в качестве гетерорегуляторных рецепторов (гетерорецепторов) в не-серотонинергических синапсах (например, в норадренергических, дофаминергических или глутаматергических), в которых постсинаптических 5-HT₁A-рецепторов нет. В этих синапсах в норме серотонин ингибирует через 5-HT₁A гетерорецепторы высвобождение других медиаторов, таких, как норадреналин, дофамин или глутамат. Агонисты 5-HT₁A-рецепторов делают то же самое вместо серотонина. Клиническое значение этого гетеро-ингибирования неизвестно, однако предполагается, что оно должно ограничивать клиническую эффективность «чистых» 5-HT₁A-агонистов и их применимость в реальной практике (а не в экспериментах на животных) в качестве антидепрессантов. Вероятно, для того, чтобы быть эффективным антидепрессантом, препарат должен, помимо 5-HT₁A-агонистической активности, обладать ещё и другими полезными для антидепрессанта свойствами (например, ингибировать обратный захват моноаминов или быть агонистом либо антагонистом ещё каких-то подтипов серотониновых рецепторов, как вилазодон или вортиоксетин). Вместе с тем, относительно «чистые» 5-HT₁A-агонисты (такие, как буспирон) или средства, обладающие среди прочих свойств 5-HT₁A-агонистической активностью (ряд атипичных антипсихотиков, особенно арипипразол, а также йохимбин, лизурид) могут быть эффективны в качестве адъюванта (дополнения) к традиционным антидепрессантам, для усиления (потенцирования) их действия и/или ускорения наступления эффекта, то есть в качестве потенцирующих агентов (аугментаторов).
Взаимодействия рецептора с другими белками
Больше 16 лет Биомолекула рассказывает о биологии и медицине — сейчас у нас на сайте несколько тысяч статей. Если вам нравится наш сайт и вы хотите, чтобы он дальше работал, поддержите нас, пожалуйста, посильной суммой — разово или ежемесячно. Ежемесячные платежи предпочтительнее 😀
Все предметы и оттенки окружающего мира мы видим благодаря сложной работе наших органов зрения. Не последняя роль в этой системе отведена рецепторам сетчатки глаза — палочкам и колбочкам.
Палочки и колбочки — что это?
Палочки и колбочки — это особые рецепторы глазного яблока, которые отвечают за передачу световой энергии и ее трансформацию в нервный импульс. Нервный импульс в свою очередь передает информацию в головной мозг, где формируется реальное изображение.
Палочки воспринимают только светлое и темное излучение, то есть только черно-белое изображение. Колбочки же распознают различные цвета и являются показателем остроты зрения. Слаженная работа рецепторов и особенность их строения обеспечивают высокую остроту зрения.
Палочки напоминают по форме цилиндр, почему и получили такое название. Их делят на четыре сегмента:
Энергия приводит палочки к возбуждению, которое человек воспринимает как свет и поэтому может видеть предметы даже при низкой освещенности. В палочках содержится особый пигмент — родопсин (основной зрительный пигмент, отвечающий за возникновение зрительного возбуждения).
Колбочки по форме напоминают — соответственно — колбочки. В них содержится другой пигмент — йодопсин, обеспечивающий восприятие зеленого, синего и красного цветов. Под воздействием света различных длин волн происходит разрушение зрительных пигментов (родопсина и йодопсина) и образование нервных импульсов, отвечающих за формирование зрительного образа.
Итак, основная функция этих рецепторов — восприятие световых волн и их трансформация в зрительный образ. Палочки помогают нам видеть в сумерках, а колбочки — при нормальном освещении.
Симптоматика заболеваний палочек и колбочек
Палочки и колбочки составляют 1 из 10 слоев сетчатки и повреждаются при ее заболеваниях. Среди основных заболеваний выделяют:
При развитии описанных патологий возникают следующие симптомы:
Подобные признаки могут сигнализировать об очень многих заболеваниях глаз, при появлении любых ухудшений зрения мы советуем обратиться сразу к врачу-офтальмологу.
Диагностика и лечение заболеваний палочек и колбочек
Для выявления заболеваний, при которых происходит поражение палочек или колбочек, доктор проводит различные исследования:
Лечение заболевания подбирается индивидуально в каждом случае и проводится комплексно: в первую очередь с устранения причины развития патологии.
Пройти полное обследование органов зрения можно в Глазной клинике доктора Беликовой. Мы используем только качественное современное оборудование и сопровождаем Пациента на всем пути — от диагностики до полного выздоровления.
Статья на конкурс «Био/Мол/Текст»: За миллионы лет эволюции млекопитающие выработали множество способов защититься от болезней. Один из них — предотвратить попадание в организм вредных веществ, а именно — реакция на горечь. Ведь что горько — то неприятно и невкусно, а что невкусно — то не едят. Горький вкус — наиболее сложный из всех известных основных вкусов (обнаружено 25 различных рецепторов, отвечающих за его восприятие), и именно он защищает от принятия токсичных веществ, вызывая врожденную реакцию отвращения у людей. Это основная роль рецепторов горького вкуса (TAS2Rs), относящихся к семейству GPCR. Удивительно, но встречаются они не только во рту, но и по всему организму: в желудке, мозге, сердце, иммунных клетках, мочеполовой и дыхательной системах. Какие же роли они там играют?
Код опасности
Эволюция горького вкуса связана с особенностями строения различных химических веществ. Историю изучения горького вкуса позволяют проследить фенилтиокарбамид (PTC) и пропилтиоурацил (PROP).

Рисунок 1. Факсимиле заголовка и первого абзаца статьи Артура Фокса о чувствительности к фенилтиокарбамиду в Proceedings of the National Academy of Sciences.
Дальнейшее развитие событий связывают с именами Рональда Фишера, Эдмунда Форда и Джулиана Хаксли, которых объединяла идея естественного отбора как движущей силы эволюции. В 1939 году они проверили, смогут ли шимпанзе попробовать воду с добавлением PTC, который некоторые люди находят тошнотворно горьким, а другие, наоборот, не ощущают горечи. История произошла в Эдинбургском зоопарке. Когда шимпанзе сделал глоток воды, он посмотрел в глаза Фишеру и плюнул в него.
И только семь десятилетий спустя был идентифицирован ген TAS2R38 (T2R38), ответственный за чувствительность к PTC. Также было подтверждено, что его различные варианты в значительной степени объясняют, почему не все могут почувствовать горький вкус фенилтиокарбамида.
Некоторые люди имеют чрезвычайно высокую чувствительность к горькому вкусу из-за высокой экспрессии генов TAS2R. Этих людей называют «супердегустаторами». Их чаще всего можно встретить в некоторых частях Азии, Африки и Южной Америки. И связано это, вероятно, с тем, что в этих районах изначально была более высокая концентрация токсичных растений по сравнению с другими континентами. Чтобы компенсировать увеличение количества потенциальных токсинов, живущие в этих районах люди могли получить больше пользы от рецепторов горького вкуса.
Каким же образом человек при относительно небольшом количестве рецепторов может ощущать горечь огромного множества химически разнообразных горьких веществ?
Как мы воспринимаем горечь
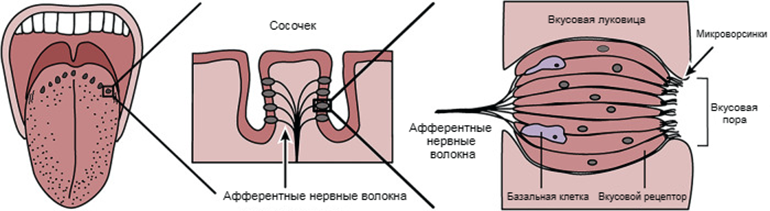
Рисунок 2. Анатомия языка. Клетки вкусовых рецепторов сгруппированы в пучки под поверхностью сосочков, где они через поры контактируют с окружающей средой.
Когда сигнальная молекула активирует рецептор, это выражается в изменении его конформации и активации G-белка. В результате его субъединицы разделяются на две части: α-субъединицу и β/γ-димер. Обе части продолжают крепиться к мембране, но не связаны более с GPCR, а потому могут свободно перемещаться и взаимодействовать с другими мембранными белками.
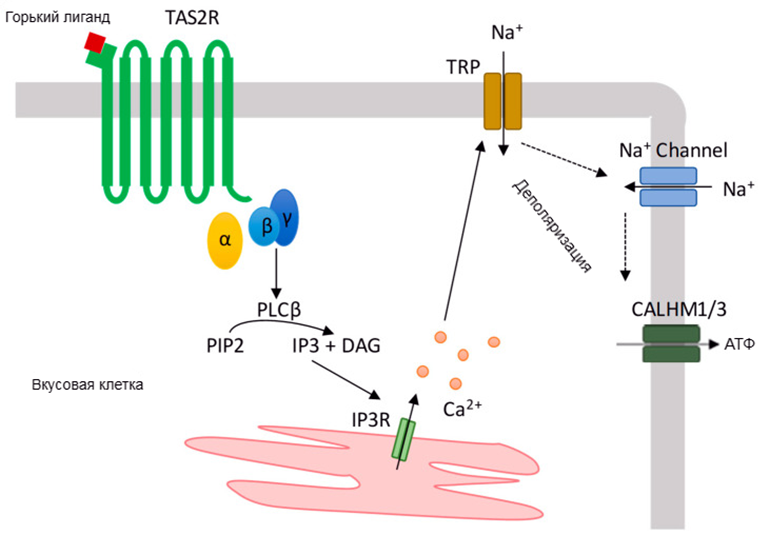
В одной и той же клетке для разных горьких веществ одновременно работает множество TAS2R-рецепторов. Эти горькие соединения действуют на одни и те же элементы внутриклеточного сигнального каскада. Возможно, именно поэтому всё горькое ощущается нами одинаково — чего, кстати, нельзя сказать о сладком. Но стоит учесть, что в повседневной жизни мы практически не сталкиваемся с чистыми горькими вкусами.
Супердегустаторы, дегустаторы и недегустаторы

Рисунок 4. Бумага, пропитанная PTC, используется для проверки, является ли человек «супердегустатором», «дегустатором» или «недегустатором».
Люди, обладающие двумя копиями PAV, сообщают, что PTC/PROP более горькие, чем гетерозиготы TAS2R38, а люди, обладающие двумя копиями AVI, вообще не чувствуют горечи PTC/PROP.
Примерно 25–30% людей — недегустаторы. Помимо того, что они менее восприимчивы к горьким вкусам, недегустаторы менее чувствительны и к сладким вкусам, терпкости алкоголя, остроте перца чили и текстуре жиров. Среди них больше курильщиков, чем среди супердегустаторов. Поэтому чувствительность к PTC/PROP часто рассматривается как общий маркер остроты вкусовых ощущений. Если вы захотите узнать, супердегустатор ли вы, то можете сделать это с помощью бумаги, пропитанной PTC (рис. 4).
Чрезмерная чувствительность к горьким веществам может быть серьезным препятствием к употреблению многих продуктов и лекарств. Но при этом быть супердегустатором — не так уж и плохо, как оказалось. Такая суперспособность таит в себе источник здоровья и долголетия.
Полиморфизмы и здоровье
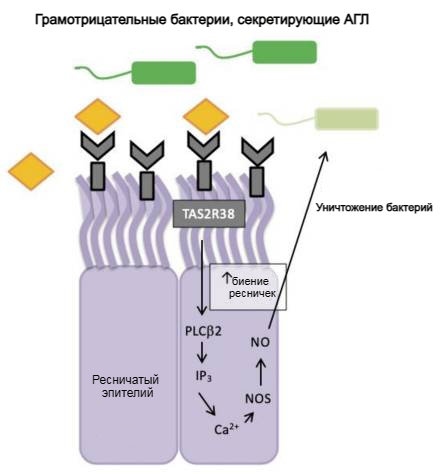
Рисунок 5. Регуляция рецептора горького вкуса TAS2R38 во врожденном иммунитете синоназального эпителия человека. АГЛ бактерий связывается с рецептором TAS2R38 и запускает каскад передачи вкусового сигнала, активируя PLC-β2 и высвобождая внутриклеточный кальций, который, в свою очередь, активирует синтазу оксида азота (NOS) и приводит к продукции оксида азота (NO). Оксид азота и его производные повреждают мембраны, ферменты и ДНК бактерий, а также увеличивают частоту биения ресничек.
До сих пор нет убедительных доказательств того, что риск развития рака зависит от потребления с пищей компонентов горького вкуса. В 1995 году диетологами-эпидемиологами из Университета Лидса было организовано когортное исследование британских женщин для изучения связи между питанием и раком. Предполагалось, что женщины с более высокой чувствительностью к горькому вкусу потребляют меньше овощей и потому чаще болеют раком. Несмотря на то, что эта гипотеза не подтвердилась, было обнаружено, что среди пожилых женщин чувствительность к горькому вкусу была связана с повышенным риском рака. У супердегустаторов риск заболеть раком был примерно на 58 процентов, а у дегустаторов — на 40 процентов выше по сравнению с женщинами-недегустаторами. При этом было отмечено, что супердегустаторы и дегустаторы ели не меньше овощей, чем недегустаторы.
Исходя из имеющихся на сегодняшний день данных, есть много оснований полагать, что рецепторы горького вкуса вовлечены в клеточные процессы рака, и это делает их ценными мишенями для разработки новых терапевтических средств.
TAS2R и пищевое поведение

Рисунок 6. Ребенок, который отказывается есть овощи.
В то время как младенцы автоматически отвергают горькие раздражители, взрослым людям проще себя контролировать, когда они ощущают горечь. Яркий пример — кофе и алкоголь. Несмотря на горький вкус, люди любят эффект, который получают от этих напитков. Но при этом многие маскируют горечь, например, разбавляя алкоголь сладкой газировкой и добавляя в кофе сахар и сливки.
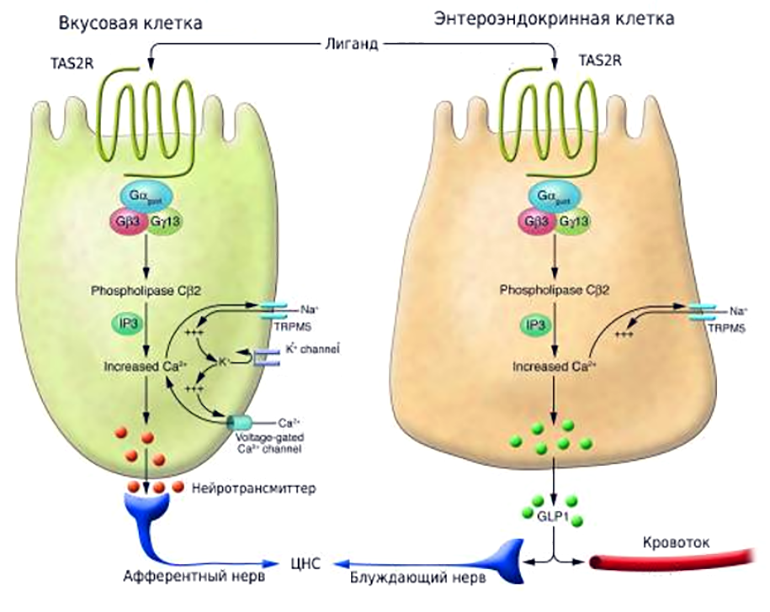
Рисунок 7. Сходство и различие в механизмах восприятия горьких веществ, рецепторами языка и энтероэндокринных клеток кишечника. И вкусовые клетки языка, и энтероэндокринные клетки (ЭЭК) имеют рецепторы TAS2R, которые обнаруживают горькие лиганды. Активированный TAS2R на ЭЭК передает сигнал внутрь клетки по пути, схожему с тем, который показан для клеток языка, и приводит к высвобождению гормонов (на рисунке — глюкагоноподобный пептид-1, GLP-1). После чего эти гормоны, с одной стороны, стремятся в кровоток и стимулируют высвобождение инсулина из клеток поджелудочной железы, а с другой — передают сигнал в центральную нервную систему (ЦНС), регулируя аппетит.
Заключение
О рецепторах горького вкуса можно написать еще много. Здесь представлена лишь часть известного о них на сегодняшний день. А сколько еще неизведанного, учитывая их разнообразие и распространение по всему организму!
Горечь многолика, а функции горьких рецепторов выходят за рамки обычного вкусового восприятия. Они регулируют самые разные защитные реакции — иммунологические, поведенческие, метаболические — и выступают потенциальными терапевтическими мишенями. Возможно, в будущем с помощью горьких веществ (агонистов рецепторов TAS2R) можно будет успешно лечить астму, хронический риносинусит, ожирение, диабет и даже рак. И смысл фразы «Хорошее лекарство горчит» станет еще более очевидным.